
INTRODUCCIÓN
Los murciélagos son los únicos mamíferos capacitados para volar (Martínez, 2011; Simmons, 2005; Varona, 1980), y constituyen el segundo orden mejor representado en la clase Mammalia, luego de Rodentia. Conforman algo más del 20 % de los mamíferos actuales, y solo no habitan la Antártida. Hasta el 2005, 1100 especies se encontraban agrupadas en 200 géneros –250 con la suma de los táxones fósiles–, con 18 familias vivientes y 6 extintas (Gunnell y Simmons, 2005; Simmons, 2005), mayormente del suborden Microchiroptera (Sevilla García, 1988, p. 116), únicos capaces de ecolocalizar (Lehel, 2006). En la actualidad (2022), según datos moleculares, se han reconocido 3 nuevas familias, y descrito más de 5 nuevos géneros, adicionalmente más de 175 nuevas especies han sido nombradas, mientras 120 especies han dejado de ser sinónimos desde 2005 (Simmons y Cirranello, 2022). Se han reportado 380 especies y 98 géneros de murciélagos neotropicales (Solary y Martínez-Arias, 2014).
Restos fosilizados se registran desde el Eoceno Temprano (51 ma), (Gunnell y Simmons, 2005). El taxon más antiguo conocido es Icaronycteris index Jepsen, 1966 [Icaronycteridae] de Norteamérica (Gunnell y Simmons, 2005; Jepsen, 1966). Análisis moleculares recientes sitúan a los protoquirópteros, posiblemente existentes a inicios del Terciario 70 Ma (Lehel, 2006), agrupados en el clado Laurasiatheria (Mancina y Borroto-Páez, 2011). Se conocen 5 géneros extintos referidos a la superfamilia Nataloidea por Simmons y Geisler (1998), no considerados miembros estrictos de Natalidae (Morgan y Czaplewski, 2003), y especies extintas en varios continentes (Mancina, 2011; Martín, 1972).
La familia Natalidae Gray, 1866, restringida al Neotrópico Americano (Morgan y Czaplewski, 2003), fue considerada monogenérica (Natalus Gray, 1838), con tres subgéneros (Mitchell, 1965; Tejedor et al., 2005), donde Natalus es el género más diverso (Tejedor, 2011; Tabla V). Según Goodwin (1959), este género agrupa tres especies, una N. major Miller,1902, es propia de las Antillas Mayores (Baker y Genoways, 1978; Griffiths y Klingener, 1988), con tres subespecies: N. m. primus Anthony, 1919 [Cuba], N. m. jamaisensis Goodwin, 1959 (Jamaica), y N. m. major Goodwin, 1959 (La Española), (Hoffman et al., 2019; Hoyt y Baker, 1980; Silva Taboada, 1974; Tejedor et al., 2004; Tejedor et al., 2005, entre otros). Cuba es centro de biodiversidad de murciélagos en el Caribe, con 27 especies vivientes (Hoffman et al., 2019; Orihuela et al., 2020), donde tres especies de natálidos son cavernícolas obligados (García Rivera y Mancina, 2011). Desde 1837, cuando por vez primera el naturalista Gervais hace mención de murciélagos hoy pertenecientes a la familia Natalidae (al estudiar materiales procedentes de una expedición a Cuba), la taxonomía de este grupo ha sido muy controvertida (Ottenwalder y Genoways, 1982).
Las especies de Natalidae tienen necesidad de vivir en cuevas, son de alimentación insectívora, con orejas en forma de embudo y presencia de órgano natálido en los machos, formado por una glándula en forma de saco situada al dorso de la base del hocico, que en Natalus es grande, con forma de campana que le cubre toda el área facial (Álvarez-Castañeda, 1999; Hoyt y Baker, 1980). Natalus primus es el mayor de los natálidos (Goodwin, 1959: Tejedor, 2011; Tejedor et al., 2005). Sus restos fosilizados han sido reportados en Ralph´s Cave, Abaco, Bahamas, con antigüedad de 3700–3570 AP (Soto-Centeno y Steadman, 2015). En Cuba se consideró extinto hasta 1992 (Walker et al., 1975), cuando fue descubierta una colonia en una cueva del occidente de la isla (García Rivera y Mancina, 2011; Tejedor et al., 2004; Tejedor et al., 2005). Su historia natural se conoce de cueva La Barca, en el occidente cubano (500 metros de espeleometría, compuesta por seis cámaras y dos cuerpos de agua), considerada una cueva neotropical caliente, con pequeñas entradas y topografía que permite una mínima ventilación, donde una de sus cámaras se caracteriza por mantener unos 26–40 °C de temperatura y una humedad relativa mayor de 90 %, características de este tipo de cueva (Tejedor et al., 2005). Las cámaras restantes son de “temperatura de fase”, donde las condiciones climáticas son intermedias; cerca del 80 % de las localidades fósiles de N. primus en Cuba y otras islas del Caribe son cuevas calientes (Tejedor et al., 2004).
En la presente contribución registramos un nuevo cráneo subfósil de Natalus primus, recobrado en la cueva de Las Anemolitas, Gibara, en el extremo oriental del archipiélago.
OBJETIVO
-Registrar la presencia de Natulus primus sobre la base de un cráneo subfósil recobrado del piso de la cueva Las Anemolitas, Gibara, Cuba.
MATERIALES Y MÉTODOS
Área de estudio. Cueva Las Anemolitas se localiza en el Municipio Gibara, al norte de la provincia de Holguín, al oriente de Cuba (760 07’ 54” de longitud oeste y los 210 06’ 34” de latitud norte), con un área de 619 km2 de extensión (Fig. 1). Sus límites son el océano Atlántico al Norte, y los municipios siguientes, dentro de la isla mayor: Holguín, al Sur; Rafael Freyre, al Este; y Calixto García al Oeste (Fernández Velázquez et al., 2014). A través de su extensión se observan tres regiones naturales que incluyen: 1) Llanura Costera (norte); 2) Sierra de Gibara (oeste), donde se destacan dos grupos montañosos: Alturas de Cupeicillo, y más al sur la Sierra de la Candelaria, con altura máxima del municipio, 243 metros en la Loma de Abelardo; 3) Llanuras Onduladas de Colinas Residuales (suroeste). Casi el 50 % de la superficie municipal está compuesta por rocas calizas, sedimentadas en mares poco profundos del Cretácico Inferior, que han permitido el desarrollo de diversos accidentes cársicos. El área cársica de la región ha sido clasificada como cónico y de torres, dentro de la región Maniabón (Núñez Jiménez, 1984), con predominio de diferentes formas: dolinas, cenotes, aguadas, lapiéz, colgadizos marinos y cuevas. La cueva ‘Las Anemolitas’ es una de estas cavidades, situada a 100 metros de altitud en el llamado Cerro Negro, localizada en el sector SO de la estribación oriental de la Sierra de Cupeicillo, a 4 km de la ciudad de Gibara (Fig. 2). Las rocas donde se abre pertenecen a la Formación Embarcadero, con edad estimada entre Paleoceno Superior-Eoceno Medio (Fernández et al., 2014). La cavidad presenta 5 entradas, y sus galerías se desarrollan en dos direcciones, NNE y SSE, su espeleometría llega a alcanzar los 630 metros de longitud.
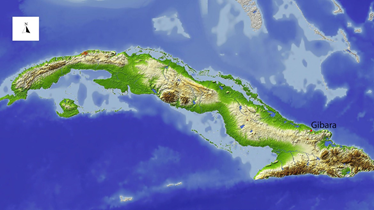
Figura 1. Ubicación de la localidad de Gibara en el nordeste cubano.
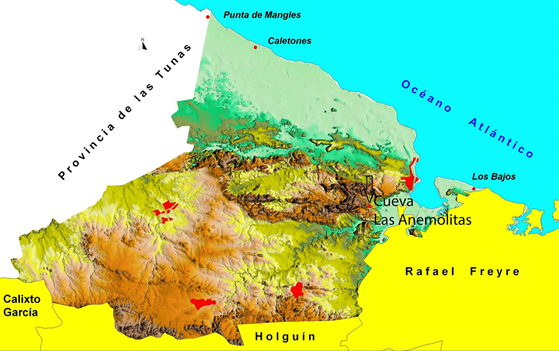
Figura 2. Situación de la cueva Las Anemolitas en el Cerro Negro, Gibara.
Materiales y ordenamiento taxonómico. Las comparaciones directas se efectuaron con material craneal de los géneros de Chiroptera presentes en la región donde se ubica el depósito: Chilonatalus Miller, 1898, y Nyctiellus Gervais, 1855, depositado en la colección del Museo de Historia Natural de Gibara “Joaquín Fernández de la Vara Pi” (MHNG 2422–2432), y las colecciones particulares del espeleólogo Alexis García Silva (ASG 398–407), y del autor (LMR–Q 674–684). Los caracteres cualitativos de niveles genérico y específico identificados son los descritos e ilustrados por Silva Taboada (1979), (clave dicotómica para los caracteres internos); Tejedor (2011) y Tejedor et al. (2005), entre otros. El arreglo sistemático sigue la lista taxonómica combinada de Vela Rodríguez y León Serafín (2007), y la actualización taxonómica de Vela Rodríguez y Silva Taboada (2020). La terminología dental es la propuesta por Duque-Osorio et al. (2009). Las observaciones del cráneo se realizaron con un estereoscopio Novel, HG661414. Las medidas (mm), se tomaron con un calibrador Vernier de precisión 0.05 mm.
RESULTADOS
Sistemática paleontológica
Clase Mammalia Linneo, 1758
Orden Chiroptera Blumembach, 1779
Suborden Microchiroptera Dobson, 1875
Superfamilia Vespertilionoidea Weber, 1928
Familia Natalidae Gray, 1866
Género Natalus Gray, 1838
El cráneo en estudio es referido al género Natalus por presentar fórmula dentaria craneal: I2, C1; PM3, M3 = 18 dientes (Silva Taboada, 1979; Walker et al., 1975). Cráneo largo y estrecho, hocico elongado con elevación marcada de su ápice y una característica acanaladura entre los huesos nasales. Longitud occipito-premaxilar mayor de 19 mm; diámetro cerebral menor de 11 mm; presencia de forámenes palatinos anteriores (bóveda palatina sin escotadura en el borde anterior); bóveda palatina no penetrada por la fosa interpterigoidea; diámetro anteorbital mayor de 3.9 mm; longitud occipito-premaxilar menor de 21.5 mm; diámetro anteorbital menor de 5.5 mm (Silva Taboada, 1979).
Natalus primus Anthony, 1919
Murciélago de Orejas de Embudo Grande
Material Referido: Cráneo [ASG–1233] con fractura del hemisferio izquierdo y escasa abrasión, colectado por Sandy Villar, en 2010 sobre un gran megalito en el piso del salón de igual nombre, cueva Las Anemolitas, municipio Gibara, provincia Holguín.
Comparaciones y descripción. El cráneo objeto del presente estudio es de color amarillento, y poco mineralizado, alargado, con caja cerebral ovalada, globosa, con cresta sagital poco desarrollada (Fig. 3A). Rostro inflado dorsoventralmente, largo (causado por la elongación rostral del premaxilar, medido desde la parte más estrecha de la constricción anteorbital hasta el ápice) en relación con la longitud total del cráneo, mayor que en las demás especies del género; formando diastema entre el canino y los incisivos –algunos autores (Sevilla García, 1988: 126), lo atribuyen a la común pérdida del I3 en murciélagos insectívoros. Ápice del rostro recurvado hacia arriba (Fig. 3B), con surco largo y profundo entre los nasales; bóveda craneana más alta en la región interparietal, lateralmente no elevada de forma abrupta sobre el plano de los huesos nasales (rostro), sino en un ángulo menor de 55o con el plano de la superficie de estos. Punto de flexión entre rostro y neurocráneo situado ligeramente más caudal, o posterior, al borde rostral (anterior) de la órbita. Constricción anteorbital con lados divergentes rostralmente; maxilar convexo (inflado) sobre los molares. Foramen infraorbitario abierto lateralmente, con un largo canal infraorbital, extendido desde P2 al M2. Ventralmente los bordes internos de las hileras dentarias son casi paralelos, con erupción total de la dentición (adulto), forámenes palatinos anteriores situados a nivel del borde anterior de los caninos, fosa basiesfenoidea dividida por un tabique. La fórmula dental craneal: I2, C1, PM3, M3 x 2, molares con patrón W marcado (Silva Taboada, 1979), tipo dilambdodonto [sin fusión de las cúspides principales externas (paracono-metacono)], adquiriendo sus ectolophos o crestas, al irradiar (labial o bucalmente) formas de W, donde la cúspide protocono queda fuera del recorrido del ectolopho, o puede estar ausente, situándose labial al paracono, formando un valle entre ambos (cuenca protoconal del Trigono), la cual recibe al Talónido del molar inferior durante la oclusión, posterior a esta se sitúan el hipocono y la cuenca hipoconal (Freeman, 1998); las tres puntas de la W serían cúspulos nombrados antero-posteriormente (parastylo, mesostylo y distostylo), una característica de la dentición de otras familias de murciélagos insectívoros como Vespertilionidae; en estos mamíferos cada molar mandibular ocluye entre dos superiores, donde los PM4 forman parte funcional de la hilera molar, allí ocluyen los molares dilambdodontos de altas cúspides (Duque-Osorio et al., 2009, Fig. 9B; Gunnell y Simmons, 2005). Fosa basiesfenoidea larga y profunda, cóncava (como Chilonatalus), forma una doble concavidad mediante una cresta medial longitudinal, con fenestra esfenoidal postorbital presente. Región interparietal más alta que la supraoccipital, con dos pares de largas fenestras palatinas entre los últimos molares maxilares (Fig. 3C).

Figura 3. Cráneo de Natalus primus recuperado en Cueva Las Anemolitas, comparado con Tejedor et al. (2005). A) Vista sagital; B) vista lateral; C) vista oclusal.
La especie Chilonatalus macer (Miller, 1914) se diferencia por presentar cráneo pequeño y alargado, con menor longitud, 13.5–14.7 mm (Kerridge y Baker, 1978; Silva Taboada,1979); rostro tan largo como la caja cerebral; fosa interpterigoidea dividida longitudinalmente por un tabique; premolares sucesivamente mayores, caja cerebral levantada de forma abrupta con relación al rostro (en ángulo dorsal a la órbita, en vista lateral), y fosas basiesfenoideas divididas en dos por cresta longitudinal con lados paralelos (semejante a Natalus, pero con cráneo mucho más pequeño) que divergen caudalmente, con un rostro más alargado desde la constricción anteorbital (Kerridge y Baker, 1982), estrecho como el cráneo, con surco entre los nasales, caja cerebral globular y delicada, elevándose desde el rostro en una curva de suave pendiente situada dorsal a la órbita (moderada flexión rostral), maxilar convexo dorsal a los molares (Tejedor, 2011, Tabla IV).
El taxon Nyctiellus lepidus (Gervais, 1837), difiere por la extremadamente pequeña longitud de su cráneo (12.5–14 mm), considerado el miembro más pequeño de la familia, y uno de los murciélagos más pequeños del mundo (Tejedor, 2011), cráneo poco inflado, caja cerebral baja, con ancho y profundo rostro, y cresta sagital muy alta, arco cigomático robusto (profundo) en relación al último molar; la emarginación palatina premaxilar y los incisivos superiores, casi se tocan entre sí a lo largo de la línea media; maxila hinchada sobre los dientes molares, con leve flexión craneal; porción anterior de los arcos cigomáticos más anchos que dos veces la altura de los últimos molares; fosas basiesfenoideas no divididas por cresta longitudinal que lo diferencia del género Natalus (Tejedor, 2011).
En N. primus (Fig. 6), existe dimorfismo sexual con variaciones morfométricas notables, el género en las Antillas mayores posee mayor tamaño que sus representantes continentales, además, las muestras fósiles de este género atribuidas al Pleistoceno son de mayor longitud, respecto a las tomadas en ejemplares vivientes en cuatro dimensiones craneales seleccionadas para el estudio: en ejemplares pleistocénicos la longitud occipito-premaxilar es 19.7–20.4 mm (Tejedor et al., 2004, Tabla III), mientras en los vivientes es solo 18.5–19.9 mm (Tejedor et al., 2004, Tabla II), valores indicativos de variación geográfica y cronológica (Tejedor et al., 2004).

Figura 6. Aspecto de Natalus primus.
Un estudio de la literatura (autores mencionados) permitió constatar caracteres cuantitativos presentes en el espécimen aquí tratado (Tabla I), los cuales separan cualquier otro natálido antillano: longitud occipito-premaxilar 18.1–19.9 (García Rivera y Mancina, 2011; Tejedor, 2011; Tejedor et al., 2005).
Tabla I. Medidas en el cráneo ASG–1233, cueva Las Anemolitas, Gibara, Cuba

DISCUSIÓN
El cráneo ASG- 1233 es referido aquí a Natalus primus Anthony, con localidad tipo Cueva de los Indios, en Daiquirí, provincia de Santiago [antiguamente Oriente] de Cuba (Anthony, 1919; Varona, 1974), ubicada a 106 km (66 millas) de distancia del presente hallazgo. Este taxon es considerado como el mayor de los natálidos (Goodwin, 1959; Tejedor, 2011; Tejedor et al., 2005). La especie se conocía por restos óseos en 22 localidades fosilíferas de Cuba e Isla de la Juventud [= Isla de Pinos] (Tejedor, 2011), hasta 1992, cuando fue descubierta una colonia en la cueva La Barca (véase Introducción), situada al occidente de la isla (García Rivera y Mancina, 2011; Tejedor et al., 2004; Tejedor et al., 2005), hallazgo que amplió la distribución conocida para este pequeño mamífero alado, actualmente en peligro crítico de extinción (García Rivera y Mancina, 2011; Mancina y Borroto-Páez, 2011; Orihuela et al., 2020). Los miles de ejemplares hallados de esta especie ocupan las cámaras calientes, colgando en grupos aislados lejos de la entrada cerca de un metro de alto, en grupos de algunas docenas a cientos de ejemplares para controlar la temperatura, sostenidos de una o ambas extremidades, sin contacto ventral con el substrato, y regularmente espaciados, a más de 10 cm (Tejedor, 2011; Tejedor et al., 2004). Convive con seis especies: Pteronotus parnellii, Mormoops blainvillei, Brachyphylla nana, Erophylla zezekorni, Artibeus jamaisensis y Chilonatalus micropus. Al parecer el N. primus realiza una suerte de migración estacional fuera de la cueva, algo improbable en otros natálidos (Tejedor et al., 2004).
La fauna viviente y extinguida de murciélagos conocida de Gibara, cuenta con 20 especies, agrupadas en 17 géneros bajo 5 familias, incluida Natalidae, representada por los géneros: Nyctiellus en 12 cuevas, Chilonatalus en 3 cuevas, conviviendo ambas sintópicamente en dos de ellas. Se han encontrado ambos géneros en estado fósil en una de ellas. En el cuaternario antillano, 27 especies de murciélagos han sufrido extinción (Tejedor et al., 2004), y el 75 % de los eventos de extinción han involucrado a murciélagos cavernarios. La casi extinción de Natalus primus parece estar dada por la dependencia a vivir perchando (colgando) dentro de las cuevas, lo cual introduce un alto grado de estoclasticidad en los murciélagos cavernarios obligados. Su limitada capacidad de dispersión debido a su vuelo lento, extrema tendencia a la deshidratación, especialización para vivir en las cada vez más escasas cuevas calientes, con frecuente colapso de secciones de estas cuevas (eventos de extirpación asociados a accidentes estoclásticos) debido a la erosión, causan el descenso de la temperatura interior alterando su ecosistema (Tejedor et al., 2004).
CONCLUSIONES
La cueva Las Anemolitas parece haber sido por algún tiempo un lugar muy caliente y húmedo, ideal para ser habitado por especies de murciélagos cavernarios obligados, como los representantes de la familia Natalidae, incluyendo el N. primus. Posteriormente, factores abióticos como el calor, la oscuridad, humedad y otros, parecen haber sido alterados por algún derrumbe que finalmente comunicó la cueva con el exterior (a través de la entrada hoy llamada El Mirador), con cambios en las condiciones interiores y causando la desaparición de la comunidad de murciélagos cavernarios local del Salón de los Megalitos (nombrado así por la gran cantidad de enormes clastos resultantes de un antiguo derrumbe). En esta cueva existió la más alta diversidad de especies de quirópteros (11) en la región que tratamos, de ellas tres vivientes, seis conocidas por restos óseos recientes y dos fosilizados. El área de la geografía gibareña, región con favorables factores naturales, gran variedad y abundancia de nichos ecológicos para ser explotados -elevado número de cuevas, extensa región forestal, y gran área con techos de tejas de construcción colonial en la ciudad cabecera, que sirven de refugio a especies insectívoras. La cueva Las Anemolitas es la espelunca de la región, con mayor representatividad de especies, donde habitan la cavidad: Macrotus waterhousei, Tadarida brasiliensis y Nyctiellus lepidus. Restos óseos recientes procedentes de esta cueva han sido identificados como Artibeus jamaisensis, Phyllops falcatum (como visitantes ocasionales o resultado de la depredación por lechuzas [=Tytonidae], Pteronotus quadridens, Brachyphylla nana, Monophyllus redmani y Phyllonycteris poeyi. Los táxones identificados en base a elementos en estado fósil incluyen a Mormoops blainvillei y la especie aquí registrada, N. primus.
AGRADECIMIENTOS
Las primeras versiones del manuscrito fueron mejoradas gracias a la revisión crítica hecha por el Dr. William Suárez. A Orlando Sánchez Tamayo y Luis Ángel Rodríguez, por su colaboración durante las comparaciones y mediciones realizadas. Sandy Villar Bermúdez por las fotos que acompañan la investigación. Carlos Suriel y revisores anónimos mejoraron sustancialmente la versión final.
REFERENCIAS
Álvarez-Castañeda, S. T. (1999). Familia Natalidae. En S. T. Álvarez-Castañeda, & J. L. Patton (Eds.), Mamíferos del Noroeste de México (67–76). Centro de Investigaciones Biológicas del noroeste.
Anthony, H. E. (1919). Mammals collected in eastern Cuba in 1917, with description of two new species. Bulletin of the American Museum of Natural History, 41, 625–643.
Baker, R. J., & Genoways, H. H. (1978). Zoogeography of Antillean Bats. Academy of Natural Sciences of Philadelphia, Special Publication, 13, 53–97.
Duque-Osorio, J. F., Ortiz Salazar, M. A., Salazar-Monsalve, L., & Mejía Pavony, C. A. (2009). Mamíferos: Evolución y Nomenclatura Dental. Revista Estomatología, 17(2), 30–44.
Fernández Velázquez, A., Sigarreta Vilches, S., Suárez Terán, S. I., Peña Rodríguez, C., Vega Torres, A., González Gutiérrez, P. A., Rodríguez, Y., Fernández Cruz, P. S., Hernández Marrero, D., Esnard Hernández, B., Urbino Rodríguez, J., Laffita Gámez, O., Córdova García, E. A., & Leyva Bermudez, O. (2014). Monografía 1: Caracterización ambiental del municipio Gibara e identificación de factores de amenaza a la flora y la fauna. Centro de Investigaciones y Servicios Tecnológicos de Holguín, (CISAT), 1–135.
Freeman, P. W. (1998). Form, Function, and Evolution in Skulls and Teeth of Bats University of Nebraska-Lincoln. Papers in Natural Resources. Paper 9. Washington, DC. Smithsonian Institution Press, 1998 (140–156).
García Rivera, L., & Mancina, C. A. (2011). Murciélagos Insectívoros (3.3). En R. Borroto-Páez y C. A. Mancina (Eds.), Mamíferos en Cuba (140–165). Impreso por UPC Print.
Goodwin, G. G. (1959). Bats of the Subgenus Natalus. American Museum Novitates, 1977, 22.
Griffiths, T. A., & Klingener, D. (1988). On the Distribution of Greater Antillean Bats. Biotropica, 20(3): 240–251.
Gunnell, G. F., & Simmons, N. B. (2005). Fossil evidence and the origin of Bats. Journal of Mammalian Evolution, 12(1–2), 209–246.
Hoffman, J. D., Kadlubar, G., Pedersen, S. C., Larsen, R. J., Larsen, P. A., Phillips, C. J., Kwiecinski, G. G., & Genoways, H. H. (2019). Predictors of Bat Species Richness within the Islands of the Caribbean Basin. Mammalogy Papers: University of Nebraska State Museum, 293.
Hoyt, R. A., & Baker, R. J. (1980). Natalus major. Mammalian Species, 130, 1–3.
Jepsen, G. L. (1966). Early Eocene Bat from Wyoming. Science, 154(3754), 1333–1339.
Kerridge, D. C., & Baker, R. L. (1978). Natalus micropus. Mammalian Species, (114), 1–3.
Lehel, B. S. (2006). Chiropteran Evolution. Where are the limits? Babes – Bolyai University.
Mancina, C. A. (2011). Introducción a los Murciélagos. En R. Borroto Paez & C. A. Mancina (Eds.), Mamíferos en Cuba (123–133). Impreso por UPC Print.
Mancina, C. A., & Borroto-Páez, R. (2011). Generalidades de los Mamíferos. En R. Borroto Paez y C. A. Mancina (Eds.), Mamíferos en Cuba (11–21). Impreso por UPC Print.
Martín, R. A. (1972). Synopsis of late Pliocene and Pleistocene Bats of North America and the Antilles. The American Midland Naturalist, 87(2), 326–335.
Martínez González, J. (2011). Protección y Desarrollo de la Fauna. Editorial Pueblo y Educación.
Mitchell, G. C. (1965). A Natural History study of the funnel-eared bat Natalus stramineus (Tesis sometida a la facultad de Department of the Zoology Master of Science. University of Arizona, USA).
Morgan, G. S., & Czaplewski, N. J. (2003). A New Bat (Chiroptera: Natalidae) from the early Miocene of Florida, with Comments on Natalid Phylogeny. Journal of Mammalogy, 84(2), 729–752.
Núñez Jiménez, A. (1984). Cuevas y Carsos. Editora Militar (EMFAR).
Orihuela, J., Viñola, L. W., & Viera, R. A. (2020). Nuevas Localidades y Registros de Murciélagos para Cuba, con énfasis en la Provincia de Matanzas. Novitates Caribaea, (15), 96–116. https://doi.org/10.33800/nc.vi15.218.
Ottenwalder, J. A., & Genoways, H. H. (1982). Systematic Review of the Antillean Bats of the Natalus micropus – complex (Chiroptera: Natalidae). Annals of the Carnegie Museum of Natural History, 51.
Sevilla García, P. (1988). Estudio paleontológico de los Quirópteros del Cuaternario Español. Paleontologia i evolució, 22, 113–233.
Silva Taboada, G. (1974). Fossil Chiroptera from Cave Deposits in Central Cuba, with Description of Two New Species (Genera Pteronotus y Mormoops) and the First West Indian Record of Mormoops megalophylla. Acta Zoologica Cracoviensia, (3), Tomo XIX. 31. V.
Silva Taboada, G. (1979). Los Murciélagos de Cuba. Editorial Academia de Ciencias, Cuba.
Simmons, N. B. (2005). An Eocene Big Bang for Bats. Perspectives. Evolution. Science, 307(5709), 527–528. https://doi.org/10.1126/science.1108871.
Simmons, N. B., & Cirranello. A. L. (2022). Bat Species of the World: A taxonomic and geographic database. https://batnames.org/about.html.
Simmons, N. B., & Geisler, J. H. (1998). Phylogenetic relationships of Icaronycteris, Archaeonycteris, Hassianycteris, and Palaeochiropteryx to extant bat lineages, with comments on the evolution of echolocation and foraging strategies in Microchiroptera. Bulletin of the American Museum of Natural History, 235, 1–182.
Solary, S., & Martínez-Arias, V. (2014). Cambios recientes en la sistemática y taxonomía de murciélagos Neotropicales (Mammalia: Chiroptera). Theria, 5(1), 167–196. https://doi. org/10. 12933/ theria- 14–180.
Soto-Centeno, J. A., & Steadman, D. W. (2015). Fossils reject climate change as the cause of extinction of Caribbean bats. Scientific Report, 5(1), 1–7. https://doi.org/10.1038/srep07971
Tejedor, A. (2005). A new species of funnel-eared bat (Natalidae: Natalus) from México. Journal of Mammalogy, 86(6), 1109–1120.
Tejedor, A. (2011). Systematics of Funnel – Eared Bats (Chiroptera – Natalidae). Bulletin of The American Museum of Natural History, 2011(353), 1–140.
Tejedor, A., Silva Taboada, G., & Rodríguez Hernández, D. (2004). Discovery of Extant Natalus major (Chiroptera: Natalidae) in Cuba. Mammalian Biology, 69(3), 153–162.
Tejedor, A., Tavares, V. D. C., & Silva Taboada, G. (2005). Revision of Extant Greater Antillean Bats of the Genus Natalus. American Museum Novitates, (3493), 1–22.
Varona, L. S. (1974). Catálogo de los Mamíferos Vivientes y Extinguidos de las Antillas. Editorial Academia de Ciencias.
Varona, L. S. (1980). Mamíferos de Cuba. Editorial Gente Nueva.
Vela Rodríguez, H., & León Serafín, L. (2007). Lista taxonómica combinada de los murciélagos cubanos. Bioespeleología. El Explorador, Periódico Digital Espeleológico. No34. 26.2.2007.
Vela Rodríguez, H., & Silva Taboada, G. (2020). Actualización Taxonómica y Distribucional de los Murciélagos de Cuba. (I). Cuba Subterránea. Sitio Web de los Espeleólogos Cubanos. Abril 10, 2020. 7 pp.
Walker, E. P., Warnick, F., Hamlet, S. E., Lange, K. I., Davis, M. A., Uible, H. E., & Wright, P. F. (1975). Genera of Recent Mammals of the World. En Mammals of the World. (Third Ed., Vol. 1). The Johns Hopkins University Press.